- 2022-08-12 发布 |
- 37.5 KB |
- 10页


申明敬告: 本站不保证该用户上传的文档完整性,不预览、不比对内容而直接下载产生的反悔问题本站不予受理。
文档介绍
从古生物学到地球生物学的跨越
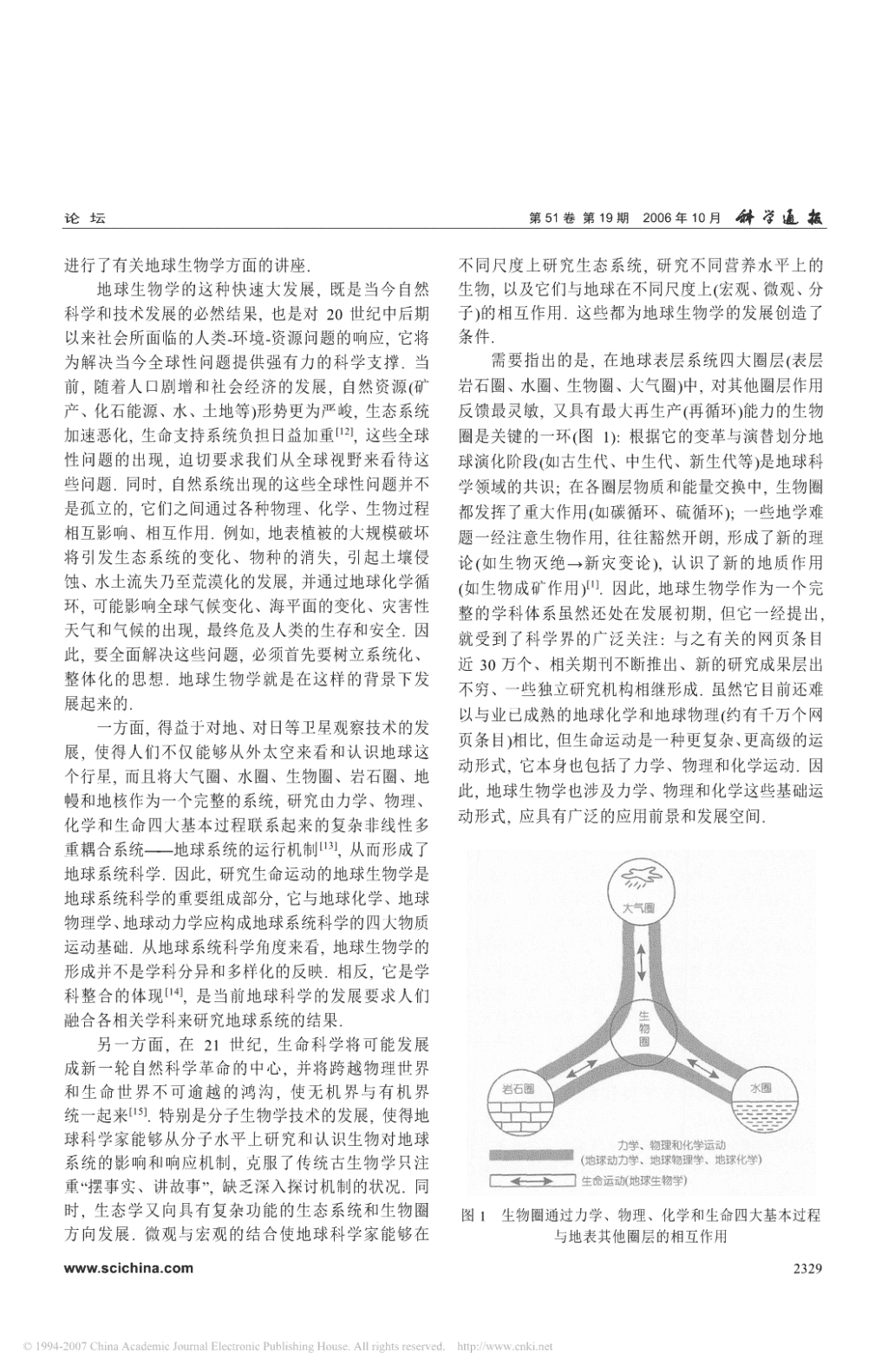
论坛第51卷第19期2006年10月从古生物学到地球生物学的跨越①②①③①②①④②谢树成龚一鸣童金南史晓颖赖旭龙Z.Q.Chen冯庆来①①①①②②王红梅杜远生王永标颜佳新张克信殷鸿福(①中国地质大学生物地质与环境地质教育部重点实验室,武汉430074;②中国地质大学地质过程与矿产资源国家重点实验室,武汉430074;③河南理工大学资源环境学院,焦作454003;④SchoolofEarth&GeographicalSciences,TheUniversityofWesternAustralia,35StirlingHighway,Crawley,WA6009,Australia.E-mail:xiecug@163.com)摘要以学科体系和科学问题为导向,对当前国际上出现的地球生物学从学科分类体系、形成背景、主要研究方向、亟待突破的分支学科及其与之相关的研究领域进行了评述.作为地球科学与生命科学相结合而形成的新兴交叉学科,地球生物学在地球科学中应具有独立的一级学科地位,类似于地球化学和地球物理学.地球生物学主要研究地球系统的生命运动,涉及地球环境与生命系统的相互作用.它的形成与发展既是当今科学技术发展的结果,也是当今世界对所面临重大人类-环境-资源问题的响应.分子地球生物学、地球微生物学、地球生态学、地球生理学等地球生物学中的二级学科还有待尽快突破,以形成地球生物学的成熟理论框架和方法体系.关键词地球生物学分子化石微生物环境生命之树协同演化中国具有得天独厚的地层古生物资源,通过几他基础学科(如数学、天文学、生物学)交叉没有形成代人的长期积累和努力探索,中国古生物学家在早独立的一级学科,而只是作为二级学科散布在其他期生命、中生代热河生物群、新生代动物辐射和生态一级学科中.例如,生物学与地球科学交叉形成的古环境变化、高精度年代地层学等领域取得了举世瞩目生物学和生物地球化学,分别放在地质学和地球化的成绩,在国际权威刊物《Nature》和《Science》上学学科中.发表相关论文数十篇,引起国际同行的广泛关注.为古生物学这一传统古老学科经历了从paleontology了巩固和发展现有的国际地位,中国古生物学家既到paleobiology,再到geobiology的发展历程.Pale-ontology着重于地质体中的古生物化石记录,突出其要发掘传统优势,更要开拓新领域.其中,学科的创地质学意义,paleobiology则是从paleontology到新和深入发展是根本.近几年,国际上出现了一系列geobiology的中间过渡环节,更加关注生物学方面的与古生物学密切相关的新术语和研究主题.在此,我意义.当前国际上出现的地球生物学(geobiology),们着重围绕地球生物学这一涉及古生物学重大变革则突出了生命系统和地球系统的相互作用.它应是问题进行分析和阐述,以促进中国相关研究领域的生物学与地球科学交叉而形成的独立一级学科.中准确定位和切入.国国家自然科学基金委员会地球科学部也建议将其1学科分类位置2)正式单独列为一个学科.在学科分类上,它等同于地球化学(geochemistry)、地球物理学(geophysics)(表地球科学(earthscience,geoscience)是人类认识、1),它们分别是研究地球物质的物理(地球物理学)、利用、改造和保护自己家园——地球的基础科学,它化学(地球化学)和生命(地球生物学)这些基础运动形是数、理、化、天、地、生六大基础自然科学之一,其1)式.由此,在地球系统中,物理、化学、生命三大基传统一级学科包括地质学、地理学、地球化学、地本过程都形成各自相对独立的学科体系(表1).球物理学、海洋科学及大气科学.这当中,只有地球有关地球生物学的定义,虽有很多,但基本雷同.化学和地球物理学是地球科学与化学、物理学两大基它强调生命系统和地球系统的相互作用,包括其机础学科交叉而形成的独立一级学科.地球科学与其[1~4]制、过程和演化历史等.生物地质学(biogeology)1)中华人民共和国教育部与中国国家自然科学基金委员会的学科分类体系有一定差异,本文的学科分类体系沿用中国国家自然科学基金委员会的体系,其一级学科相当于中华人民共和国国家标准学科分类与代码(GB/T13745-92)的二级学科2)刘羽,姚玉鹏,陈淮.地球生物学——充满挑战的新领域.国家自然科学基金委员会情况交流,2005-09-22www.scichina.com2327\n第51卷第19期2006年10月论坛[1,5]主要是利用生物学的知识来服务于地质学,是地另外,国际上出现的生物地貌学(biogeomor-[8]球生物学的一部分.近年出现的biogeosciences(生物phology)主要研究地貌与生物之间的相互影响,可地学)主要研究生命(包括其他行星上的生命)与地球以作为生物地质学(生物对地貌的作用)和地球生态[6]的大气圈、水圈和岩石圈的相互作用,它更强调将学(地貌对生物的影响)的一部分内容.生物学方法与地球物理和地球化学的方法联合起来2形成背景1)研究地球系统,而且还涉及太空生物学(astrobiology当今出现的地球生物学思想,其酝酿和萌芽阶——应是“生”与“天”两大基础学科交叉结合而形成的).[9]段可以追溯到两个世纪以前Hutton对地球表层的作为地球科学领域的一级学科,地球生物学将研究,那时已经认识到生物对地球的作用,但同时也来有可能发展出较多的二级学科,但目前的体系结认为生命系统对地球系统动力学过程所起的作用很构还很不完善(表1).总体上来说,表中所列的这些二[10]小.1934年,荷兰生物学家BassBecking开始使用级学科可以归纳为如下3类:“地球生物学(geobiology)”这个术语.随后,Love-(1)分子地球生物学(moleculargeobiology,包括[11]lock从地球生理学角度,把地球作为一个有机体,分子古生物学、分子地层学、分子生态学、分子考古研究生命系统对地球表层环境的影响.从此,人们开学等)、地球微生物学(geomicrobiology)和传统古生物始突出了地球生物学的研究主题,即强调生物圈与学,主要着眼于不同尺度(分子、微观、宏观等)的不2)同对象.地圈的相互作用,或生物界与非生物界的相互作用,[12](2)地球生态学(geoecology,包括古生态学、生并很快形成了一个全新的研究领域:2000年,在美态地层学、生态地质学、古生物地理学)和地球生理国微生物科学院召开的讨论会上正式把地球生物学描学(geophysiology),主要强调生物与环境的相互作用.述为“试图了解生物圈与地圈相互作用的研究”;2001(3)生物地球化学、生物地球物理学(如生物磁学)年,美国地质学会成立了地球生物学分会;Elsevier和生物地质学(包括生物地层学、生物成岩成矿学等[1]),和Blackwell出版社则分别于2002,2003年推出了刊侧重于研究生物对地球系统的作用.它们是地球科物《VirtualJournalofGeobiology》和《Geobiology》;学各一级学科之间交叉结合的产物,即分别是地球2004年,美国国家科学基金会地球科学部正式将地球生物学与地球化学、地球物理学、地质学相互交叉结生物学单独列为一个项目组(program),予以稳定的支合的产物.其中的生物地球化学分类位置一直放在持;同时,南加洲大学Wrigley环境研究所则开办了[7]地球化学学科中.最近,Cutter已提出,将生物地球地球生物学的国际培训班;2005年,中国国家自然科化学作为地球生物学这个学科的一个重要组成部分.学基金委员会地球科学部邀请著名学者DerekBriggs表1地球生物学与其他学科的关系及其二级学科一级学科二级学科(建议)研究对象地质学(固体地球)性质海洋科学(液)大气科学(气)略地理学(地球表层)地球物质运动地球物理学(物理运动)科形式地球化学(化学运动)学地球生物学(生命运动)分子地球生物学(分子古生物学、分子地层学、分子生态学、分子考古学、古生物化学)、地球微生物学、古生物学(古动物学、古植物学、演化生物学等);地球生态学(古生态学、生态地质学、生态动力学、生物地理学)、地球生理学;生物地球化学、生物地质学(生物地层学、生态地层学、生物成岩成矿学、生物矿物学、生物地貌学)、生物地球物理学(生物磁学等)等1)HedinL,ChadwickO,SchimelJ,etal.LinkingEcologicalBiology&Geoscience:ChallengesforTerrestrialEnvironmentalScience.WhitePaperreporttotheNationalScienceFoundation,2002.182)NealsonK,GhiorseWA.Geobiology:ExploringtheInterfacebetweentheBiosphereandtheGeosphere.AmericanSocietyforMicrobiologyReport,2001.232328www.scichina.com\n论坛第51卷第19期2006年10月进行了有关地球生物学方面的讲座.不同尺度上研究生态系统,研究不同营养水平上的地球生物学的这种快速大发展,既是当今自然生物,以及它们与地球在不同尺度上(宏观、微观、分科学和技术发展的必然结果,也是对20世纪中后期子)的相互作用.这些都为地球生物学的发展创造了以来社会所面临的人类-环境-资源问题的响应,它将条件.为解决当今全球性问题提供强有力的科学支撑.当需要指出的是,在地球表层系统四大圈层(表层前,随着人口剧增和社会经济的发展,自然资源(矿岩石圈、水圈、生物圈、大气圈)中,对其他圈层作用产、化石能源、水、土地等)形势更为严峻,生态系统反馈最灵敏,又具有最大再生产(再循环)能力的生物[12]加速恶化,生命支持系统负担日益加重,这些全球圈是关键的一环(图1):根据它的变革与演替划分地性问题的出现,迫切要求我们从全球视野来看待这球演化阶段(如古生代、中生代、新生代等)是地球科些问题.同时,自然系统出现的这些全球性问题并不学领域的共识;在各圈层物质和能量交换中,生物圈是孤立的,它们之间通过各种物理、化学、生物过程都发挥了重大作用(如碳循环、硫循环);一些地学难相互影响、相互作用.例如,地表植被的大规模破坏题一经注意生物作用,往往豁然开朗,形成了新的理将引发生态系统的变化、物种的消失,引起土壤侵论(如生物灭绝→新灾变论),认识了新的地质作用蚀、水土流失乃至荒漠化的发展,并通过地球化学循[1](如生物成矿作用).因此,地球生物学作为一个完环,可能影响全球气候变化、海平面的变化、灾害性整的学科体系虽然还处在发展初期,但它一经提出,天气和气候的出现,最终危及人类的生存和安全.因就受到了科学界的广泛关注:与之有关的网页条目此,要全面解决这些问题,必须首先要树立系统化、近30万个、相关期刊不断推出、新的研究成果层出整体化的思想.地球生物学就是在这样的背景下发不穷、一些独立研究机构相继形成.虽然它目前还难展起来的.以与业已成熟的地球化学和地球物理(约有千万个网一方面,得益于对地、对日等卫星观察技术的发页条目)相比,但生命运动是一种更复杂、更高级的运展,使得人们不仅能够从外太空来看和认识地球这动形式,它本身也包括了力学、物理和化学运动.因个行星,而且将大气圈、水圈、生物圈、岩石圈、地此,地球生物学也涉及力学、物理和化学这些基础运幔和地核作为一个完整的系统,研究由力学、物理、动形式,应具有广泛的应用前景和发展空间.化学和生命四大基本过程联系起来的复杂非线性多[13]重耦合系统——地球系统的运行机制,从而形成了地球系统科学.因此,研究生命运动的地球生物学是地球系统科学的重要组成部分,它与地球化学、地球物理学、地球动力学应构成地球系统科学的四大物质运动基础.从地球系统科学角度来看,地球生物学的形成并不是学科分异和多样化的反映.相反,它是学[14]科整合的体现,是当前地球科学的发展要求人们融合各相关学科来研究地球系统的结果.另一方面,在21世纪,生命科学将可能发展成新一轮自然科学革命的中心,并将跨越物理世界和生命世界不可逾越的鸿沟,使无机界与有机界[15]统一起来.特别是分子生物学技术的发展,使得地球科学家能够从分子水平上研究和认识生物对地球系统的影响和响应机制,克服了传统古生物学只注重“摆事实、讲故事”,缺乏深入探讨机制的状况.同时,生态学又向具有复杂功能的生态系统和生物圈图1生物圈通过力学、物理、化学和生命四大基本过程方向发展.微观与宏观的结合使地球科学家能够在与地表其他圈层的相互作用www.scichina.com2329\n第51卷第19期2006年10月论坛3主要研究方向4.1分子地球生物学(moleculargeobiology)——追溯生命之树和全球环境的分子记录20世纪后期以来,Plenum出版社出版了一系列分子地球生物学是美国国家研究理事会NRC地球生物学主题丛书,如水生生物的骨骼生长、动物(2000)创用的词,意指分子生物学、分子古生物学和与沉积物的关系、化石记录中捕食与被捕食生物间的[1]地球科学相结合的研究.研究对象是生物分子(核相互作用、地球生物圈中的碳,新元古代的地球生物酸、蛋白质和氨基酸、类脂物、纤维素等).它对应于学与化石生物学等.2003年由Blackwell出版社推出生命科学领域的分子生物学.主要研究重点有:的期刊《Geobiology》列出了9个主要领域,它与耶(ⅰ)生命起源和生物演化的分子记录鲁大学生物圈研究所所长DerekBriggs在中国国家自1)包括地学记录和生物学记录两个方面.然科学基金委员会的交流中提出的9个主要研究方地学记录——分子化石:通过对矿物表面有机反向一致.它们分别是:(1)生命的起源和演化;(2)大应的研究,可以了解地球历史早期生物出现之前有气圈、水圈和生物圈的演化;(3)地球演化关键转折机化合物是如何形成的,为生命起源提供可能的线期沉积岩记录和地球生物学;(4)古生物学和演化生索.通过沉积岩中地质类脂物分子化石及其同位素态学;(5)生物标志物;(6)分子生态学和谱系演化;[16]组成特征的研究,可能追溯生命演化中重要新陈(7)环境微生物学;(8)生物地球化学和全球元素循[17~20]代谢过程出现的时间、顺序及其作用.例如,分环;(9)微生物与矿物的相互作用.子证据表明,至少在27亿年前,具有复杂结构的真实际上,概括以上的主题和方向,地球生物学主[18]核细胞生物就可能已经出现,这比已知的最古老要研究两个最基本的方面.一是地球各圈层对生物的真核化石早5~6亿年.根据地质类脂物分子化石,圈的影响和作用,包括以上的第(1)~(6)项内容,涉及[21]有可能建立生物标志化合物的生命之树,以验证的学科主要是传统古生物学领域的一些学科,如演某些生物学记录(如校准分子钟)等.化生物学、生物地层学、生态地层学、生物地理学.这当然,太古代和元古代地质类脂物分子研究所方面的工作不仅是为了理解地球上生命的起源和演[18]面临的最大挑战是污染问题.尽管Brocks等人想化,而且也为研究太空生物学服务.二是生物圈对地尽许多办法来克服污染,但其结果仍然被一些学者球其他圈层的作用和影响,包括以上的第(7)~(9)项[22]认为可能是污染造成的,看来这方面的工作还有内容,涉及的学科主要散布在非古生物学领域的一些地质学科中,如生物地球化学(地球化学)、生物成待更多资料的积累.矿学(矿床学)、生物成岩作用(沉积学、岩石学)等.这生物学记录——分子钟和分子演化树:由于相当个方向的研究工作是为了更加深刻地理解全球规模一部分蛋白质分子进化速度具有恒定性,可以根据的环境变化,以及为预测未来的变化服务.因此,从生物DNA之间的差异,对分子演化树的各分支标定[23]古生物学跨越到地球生物学,不仅仅是从地球科学时间,建立起分子钟.大多数动物的核心进化基因的二级学科拓展到地球科学的一级学科,更重要的是相同的,植物亦是如此.根据生物分类单位的基因是其研究内容在广度和深度上均发生了实质性的变相似性,可建立起包含全部生物门类的分子演化化:更加关注生物圈在不同尺度上对地球系统的作[24]树.用;在传统古生物学注重记录的基础上,更加关注作当前的重点是如何将地学记录与生物学记录结用的过程和机制.合.演化树、分子钟与年代地质学相结合,并与化石4有待突破的分支学科和相关领域记录中新类别的出现年代相检验,可建立起包含演[25]化树各主要分支事件的地质年代表.从最早的多地球生物学作为一个备受人们关注的新兴学科,细胞动物出现到脊椎动物各纲的分化,都可以一一有待突破的分支学科和研究领域很多.在此重点讨标识.地质化石和古DNA数据可用来校准分子钟;论在为地球系统科学服务中,地球生物学需突破的分子钟也可以帮助建立其他缺乏丰富化石记录的系一些重要方面.[26]统谱系.古DNA和地质类脂物的结合已经开始.1)同第2327页脚注2)2330www.scichina.com\n论坛第51卷第19期2006年10月[27~30]中国学者在早期生命过程和中生代热河生物当前人们关注的关键科学问题有:不同背景条件下[31~35]群等领域积累了大量高水平的国际成果,为校微生物群落维持的机制,包括其能量来源、生长方式准分子钟提供了重要的地质学和古生物学依据.等;微生物的多样性;微生物之间的相互作用;微生(ⅱ)地球环境变化的分子标志物与宏体生物之间的相互作用等.例如,在富有机质作为地质历史时期生物有机体遗留下来的有机沉积物附近的砂岩中(如白垩纪砂页岩中),一些异氧分子,特别是其中具有生物学意义的分子化石,能精厌氧细菌主要靠有机物提供能量;在火成岩缺少有细记录各种不同时间分辨率的全球气候、环境、植被机物的地方,高分异度的微生物主要以H2作为能量[56]变化信息.当前研究载体已涉及全球变化的诸多关来源;而深海沉积物中的微生物群落主要依靠厌键载体,如海相沉积物[36~38]、湖泊和泥炭沉积物[39]、氧微生物的新陈代谢和硫酸盐还原作用.当前主要[40][41][42]关注的研究领域有:雪冰和冰芯、黄土-古土壤系列、洞穴石笋等.(ⅰ)微生物的起源和演化比较典型的例子有海洋沉积物长链烯酮的不饱和度[43,44]有关微生物分子演化树各节点出现的时间还远与海水表面温度的定量关系;全球性古火灾(如[45]未查明,目前备受人们关注的是蓝细菌、硫酸盐还原古近纪/白垩纪)的分子标志等.菌、甲烷菌等强烈影响地球其他圈层的微生物.生命当前,人类活动加剧了各类地球环境的变化.在科学家主要从分子钟来推算.地球科学家主要从微当代环境污染研究中,已明确提出了环境污染的化[57][58][46]生物的形态学、沉积组构(如叠层石)、碳和硫同石生物标志物和人类活动标志物.前者指化石燃[59][18]位素、分子化石等开展研究.就形态学来说,有料和沉积物中的分子化石造成的现代环境污染,最两个问题需慎重:一是如何把微生物个体与物理、化典型的例子是海洋中石油泄漏造成的污染.后者指学作用形成的有机体区分开;二是如何进一步从形人类在利用这些分子化石过程中所引起的环境污染,如人工合成的DDT,PCB(多氯联苯),PAH(多环芳烃)态上把生物学工作提升到生理学上,以便区分不同[60]等的有机污染.这些有机污染物已影响到了大气圈、生理功能的微生物,这对地球生物学的研究至关水圈(冰冻圈)、生物圈和土壤圈,有关这些污染源的重要.例如,一般细菌与营光合作用蓝细菌的区分,形成过程和消除都离不开对分子化石本身地球化学可以探讨产氧光合作用问题.值得注意的是,同位[58][18]性质的研究.另外,这些环境污染物通过食物链对不素和分子化石方面的重要进展,不仅对不同微同生物的影响过程和机理,也是分子地球生物学和生物的起源提供重要线索,也为生命早期的微生物生态学的一项研究内容.当然,这些工作也都是环境地球化学过程研究提供了切入点.值得注意的是,作科学、地球化学等学科的重要研究内容.为当代海洋主要生产力的三大微生物类群(颗石藻、[61]硅藻、沟鞭藻),其起源的时间和机制也还不完全4.2地球微生物学(geomicrobiology)——探索全球清楚,它们的起源与环境条件的关系更是个谜.变化过程中生态系统底层的状况和作用(ⅱ)全球生物危机期间的微生物状况和作用在国际上,传统古生物学的研究工作主要集中已引起人们关注的是,在一些生物集群灭绝期在真核生物方面.例如,显生宙5次生物集群灭绝事或期后,出现了钙质微生物岩(microbialite)或微生物件主要是根据地层中的真核生物(主要是动物化石)的繁盛,如志留纪/奥陶纪、晚泥盆世弗拉期/法门期、得出的结论.如果从生态系统来考虑的话,过去的工三叠纪/二叠纪界线等.这些微生物岩为研究地球微作仅涉及到生态系统上层的一小部分成员,目前还生物生态系统提供了窗口.钙质微生物岩主要由各缺乏对生态系统底层的大部分成员的系统研究工作.种微生物席(microbialmats)捕获沉积物或直接使矿如果从现行的生命构成来看,在古细菌、真细菌和真核生物三大类群中,过去的工作几乎没有或很少涉物沉淀而形成,这类岩石在前寒武纪时期曾经很常及前两大类.但恰恰是这些地球微生物,对全球变化见,多样性高.然而,随着寒武纪多细胞生物的辐射以及地球表层系统起着重要乃至关键的作用,是地演化,动物对微生物席的破坏使钙质微生物岩在显[47~53]生宙剧减[62].很有意义的是,在显生宙重大地史转折球生命的支持系统.因此,要对地质历史时期生态系统和全球变化做全面的了解,必须开展生态系期,在一些动物集群灭绝的同时或期后,这些钙质微[54,55][63,64][65]统底层——地球微生物(也称地微生物)的研究.生物岩又大量出现,如早三叠世、晚泥盆世、www.scichina.com2331\n第51卷第19期2006年10月论坛[62]晚奥陶世等.这些钙质微生物岩被认为是环境变态学强调当代生物圈(特别是人类)与作为其生存环[66]化的标志,对地史重大转折期生物危机发生时的境的地球系统的相互作用,以及对未来的预测及对[85]特殊生态系统的研究具有重要意义.因此,与动物集策.其中,人类干预下的水资源、土地资源等特别[11]群灭绝相伴的这些微生物岩受到人们的关注,它们受到地球生态学家的关注.Lovelock较早使用地球是研究地球微生物的重要载体.生理学这个术语,把地球看成是由生物界和物理界[86](ⅲ)微生物对全球气候和环境变化的响应和反馈紧密耦合的系统进行研究,特别强调把地球当作由不同生物组成的生态系统对全球或区域气候一个有机体,研究这个有机体中的生物与地球的相[87~89]环境变化的响应和反馈并非千篇一律,而在时间、空互作用.地球生态学和地球生理学就是着眼于生物与地间和强度上都存在一定的差异.仅就强度而言,地球球环境的相互作用.地球自从有了生命,生物与环境微生物对突变事件的响应可能与其他生物有所不同.就成为一对互相制约、互相促进的共同体.生物界的在陆地生态系统中,微生物对突变气候的响应可能[90]每一次突变都与地球环境的重大变化密切相关.与植被不同.地球微生物对气候、环境变化的响应直例如,新元古代后期多细胞生物群的繁盛,是在地球接关系到陆地风化作用、碳循环等一系列全球性的重[67]从冰冻的雪球状态解冻之后;寒武纪初的生命大爆大地球科学问题,进而又反馈气候环境的变化.目[68~70]发,其先决条件之一是当时大气与海洋具备了足够前,植被与气候变化的相关性备受人们的关注,的氧气;古生代、中生代之交剧烈的全球变化与生物而有关微生物与气候、环境变化的相互关系目前还知大灭绝同时发生;中生代、新生代之交恐龙的灭绝,之甚少.则与外星体撞击造成的环境剧变有关.(ⅳ)极端环境微生物在生物与地球环境的相互作用、协同演化过程中,近年来已取得突出的进展.随着大洋钻探研究当前研究比较薄弱的环节是生物对地球系统的作计划、冰川和极地的科学考察以及大陆超深钻的实施,[60]用:生物的风化侵蚀作用对水圈、土壤圈的影特别是由于微生物学家与地质学家联合发展了能避[91,92][93][71]响,生物活动对大气圈的影响,生物沉积和成免外部微生物污染的采样方法,各种极端环境下[94,95][56,72,73]岩作用对岩石圈的影响,等等.例如,一些微生微生物的陆续发现,使生物圈的范畴在不断扩[54][74,75][76,77]物利用金属元素来获得能量,就像人类需要氧气一大,由空中到地下、由冰川到热泉喷口、[78][79~81]样.这方面工作的深入必然涉及其机制问题,它是地由深海和深湖到深部地壳,等等.极端环境微球生理学的重要工作.因为微生物在其中起了关键生物正在以崭新的面貌诠释着生命的含义,同时也改[96]作用,它又是地球微生物学的研究内容.而机制的变了人们的思维方式.了解极端环境下的微生物种类,深入有待于分子水平上工作的突破,例如对金属元有助于地质学家更加清晰地认识极端环境下的地质和1)素起氧化或还原作用的微生物某种基因的筛选等,地球化学过程.极端环境微生物的研究也为生物学家所以它又是分子地球生物学的内容.因此,生物对地探索生命的起源、发现新的微生物代谢途径和生存对[54]球系统的作用需要这些二级学科的联合攻关才能深策等提供了前所未有的机遇.例如:利用有机大分[50]入,这可能也是这方面研究相对滞后的原因之一.当子(干酪根)的微生物代谢作用;利用H2作为能源的[56][49]前,这方面的一些工作已进行了一些定量化的数学微生物;厌氧环境微生物对甲烷的氧化作用,一[82]模拟工作,如生物对碳循环的影响,体现了生物学、些新的微生物的发现和分离,等等.地球科学和数学这三大基础学科的结合.4.3地球生态学(geoecology)和地球生理学(geophy-在地球生理学研究中,值得注意的是生物膜siology)——诠释生物与环境协同演化的机制和过程[97~99](biofilms)的工作,生物膜中的微生物重量是所[83]地球生态学由地貌学家Troll提出,起初用来有宏体生物的1000倍.一些生物对矿物的作用是通描述自然地理学和生态学的联系,主要研究生态系[98]过生物膜起作用的.全球板块构造的形成、地壳的统的结构和功能,重视环境中生物与非生物因子之[100,101]演化都直接或间接地与生物膜作用有关.生物[84]间的复杂作用.至今,发展相对比较成熟的地球生1)Cervini-SilvaJ,FakraS,GilbertB,etal.AmolecularapproachtowardsunderstandingthebiogenicformationofCeO2anditsinteractionswithbiomolecules.AbstractofAGUFallMeeting,2005.1262332www.scichina.com\n论坛第51卷第19期2006年10月膜的作用还使一些浮游生物能发生自沉积作用域的重大进展,例如,基因水平转移、生命之树(tree(self-sedimentation),在地质体中形成一些富有机质oflife)或生命之环(ringoflife)以及极端环境的生命纹层.它一方面避免了动物对它的捕食和破坏,另一系统等,将为地球生物学在相关领域的突破创造条方面通过自沉积作用改变生物地球化学循环、生态动件.力学结构等.因此,地质历史时期生物膜以及与之相当然,更高层次上的多学科交叉联合也将促使关的自沉积作用的识别对全球重大地质事件的研究地球生物学更深入的发展,如生命科学、地球科学与[102]具有重要意义.当前,生命科学领域已经对生物数学这三大基础学科的结合将促进地球生物学的定[103,104]膜开展了广泛的研究,但在地球科学领域,这量化研究,这些工作也才刚刚开始,这种大学科的联才刚刚开始.合还需更长时间的积累.另外,在地球环境演化过程中,生物多样性的变5.2地球生物学和地球化学、地球物理学的联合化、生态动力学等问题正引起人们广泛关注.在现代地球生物学要为地球系统科学服务,需要与地生命科学领域进行了大量的生物多样性和生态动力[7,109~112]球化学、地球物理学这些一级学科联合.而且,学工作,但在地质历史时期,除了几大生物集群灭绝生命运动本身包括力学、物理和化学运动,地球生物[105]外,有关生物多样性的变化都是一些粗线条的工学又必须与地球动力学、地球物理学和地球化学联合.作,其生态动力学原因还知之甚少.例如,在化石记地球生物学所要研究的生物与地球环境协同演化,录中,在不同环境背景、不同生物多样性条件下,作其研究深度、广度绝非古生物学这一单一学科所能解为生物与生物之间相互作用重要形式之一的捕食者[60]决的.就拿产氧光合作用来说,传统古生物学主要与被捕食者的关系(predator-prey)及其对生态系统和关注与产氧光合作用有关的蓝细菌是如何和何时起[106]生物演化的影响,虽已引起注意,但还需系统和源的,地质体中最早的蓝细菌记录是什么(形态学、沉深入.积构造、同位素或分子化石)等问题.而地球生物学则5面临的挑战更加关注在产氧光合作用出现以后,大气圈和海洋由地球科学和生命科学交叉所形成的地球生物的氧化-还原状态是如何发生改变的,它需要在传统学给传统古生物学的发展提供了机遇,同时也给传古生物学研究的基础上,结合其他能够反映大气圈统古生物学的研究带来了巨大的挑战[107].和海洋氧化-还原状态的地质学、地球化学和地球物理学记录,这样才能探讨生物圈对地球其他圈层的5.1传统古生物学与生命科学的联合与突破作用、影响和反馈.如前所述,传统古生物学关注地质记录,即注重与地球化学、地球物理学相比,地球生物学的理“摆事实、讲故事”,但“故事”中的道理难以讲透,即论框架还未真正建立.如何跳出传统研究的思想框对机制问题的研究不够深入.现代生命科学却非常架,从地球系统科学的视野来研究地球生物学,还需关注机制问题,是一个能够讲透“道理”的学科.另一要一个较长期的积累过程.当前地球动力学的发展,方面,地质历史上生命系统的研究可以为现代生命也将进一步促进地球生物学的动力学过程研究.科学的研究提供历史事实、演化记录等信息.探讨生物与环境协同演化的问题则必须建立在生命科学与5.3形态学与分子水平的结合地球科学的联合上,尽管生命科学可以依据分子钟现代分子生物学的发展成了生命科学的主流.建立分子演化树,但地质历史时期环境条件的变化目前涌现出的大量研究成果表明,分子生物学对生必须依靠地学记录来解决.因此,如何整合地球科学命科学的意义就像了解原子结构对物理学一样重要.[108]和生命科学领域的成熟技术和方法,实现地球生虽然地球科学家也正在开展分子古生物学(古DNA,物学与生命科学的真正交叉、融合,需要两个领域的地质类脂物等)的工作,但要将传统形态学工作与分科学家不断进行自觉的碰撞.这些都需要时间和过子水平工作相结合,还需要时间.即使在形态学方面,程,毕竟地球科学和生命科学这两个领域的研究在虽然积累了大量资料,但许多古生物资料是零散的.时间尺度上存在巨大差异,它们有着不同的目标,其如何集成传统古生物学的资料,为地球生物学的发[14]研究方法也很少有雷同.特别是,当前生命科学领展服务,是迫切需要我们解决的问题.在分子古生物www.scichina.com2333\n第51卷第19期2006年10月论坛学内部,古DNA和地质类脂物的结合也才刚刚开前缘,2002,9(1):11―18[25]14NoffkeN.Geobiology——aholisticscientificdiscipline.Palaeo-始.geogrPalaeoclimatolPalaeoecol,2005,219:1—3致谢感谢老一辈古生物学家在开拓和发展中国地球生物15毕思文.地球系统科学.北京:科学出版社,2003.38916王红梅,谢树成,赖旭龙,等.分子地质微生物学研究方法述评.学各分支学科所付出的艰辛努力,他们在引领中国古生物地球科学进展,2005,20(6):664670学的国际地位、培养国际水平的人才队伍方面做出了突出17SummonsRE,JahnkeLL,HopeJM,etal.2-Methylhopanoidsas的贡献,这些工作为形成中国地球生物学领域的完整学科biomarkersforcyanobacterialoxygenicphotosynthesis.Nature,体系奠定了坚实的基础.感谢中国国家自然科学基金委员1999,400:554―557会地球科学部在地球生物学领域所起的引领和导向作用.18BrocksJJ,LoganGA,BuickR,etal.Archeanmolecularfossils本文思想(如生命运动与地球生物学及其学科分类位置等)andtheearlyriseofeukaryotes.Science,1999,285:10331036得益于与张本仁院士(化学运动与地球化学)、金振民院士19OlcottAN,SessionsA,CorsettiFA,etal.BiomarkerevidenceforphotosynthesisduringNeoproterozoicglaciation.Science,(力学、物理学运动与地球动力学、地球物理学)的多次交2005,310:471—474流与深入讨论以及中国地质大学(武汉)地球生物学系所进20KnollAH.Anewmolecularwindowonearlylife.Science,1999,行的多次学科建设研讨会.感谢两位审稿专家对本文提出285:1025—1026的建设性意见.本文受中国国家杰出青年科学基金(批准21BrocksJJ,PearsonA.BuildingtheBiomarkerTreeofLife.Rev号:40525008)、中国国家自然科学基金重点项目(批准号:MineralGeochem,2005,59:233—25840232025)和中国国家自然科学基金创新研究群体项目资22KoppRE,KirschvinkJL,HilburnIA,etal.ThePaleoprotero-助.zoicsnowballEarth:Aclimatedisastertriggeredbytheevolutionofoxygenicphotosynthesis.ProcNatlAcadSci,2005,102:参考文献11131―1113623CooperA,GrasslyN,RambautA.Usingmoleculardatatoesti-1殷鸿福,杨逢清,谢树成,等.生物地质学.武汉:湖北科学技matedivergencetimes.In:BriggsDEG,CrowtherPR,eds.Pa-术出版社,2004.264laeobiologyⅡ.Oxford:BlackwellPublishing,2001.532—5342PennisiE.Geobiologists:Asdiverseasthebugstheystudy.Sci-24WangYC,KumarS,BlairHS.Divergencetimeestimatesfortheence,2002,296:1058―1060earlyhistoryofanimalphylaandoriginofplants,animalsand3KnollAH,HayesJM.Geobiology:Articulatingaconcept.In:fungi.ProcRSocLondB,1999,266:163—171LaneRH,LippsJ,SteiningerFF,etal,eds.Paleontologyinthe25AyalaFJ,RzhestkyA.Originofthemetazoaphyla:molecular21stCentury:Frankfurt,InternationalSenckenbergConference.clocksconfirmpaleontologicalestimates.ProcNatlAcadSci,KleineSenckenberg,1997,25:105—1081998,95:605—6114KnollAH,HayesJM.Geobiology:ProblemsandProspects.In:26CoolenMJL,HopmansEC,RijpstraWIC,etal.EvolutionofLaneRH,SteiningerFF,KaeslerRL,etal,eds.FossilsandthethemethanecycleinAceLake(Antarctica)duringtheHolocene:Future:Paleontologyinthe21stCentury.Senckenberg-Buch,2000,responseofmethanogensandmethanotrophstoenvironmental74:149change.OrgGeochem,2004,35:1151—11675殷鸿福.生物地质学.地球科学进展,1994,9(6):79—8227ZhuM,YuX.Aprimitivefishclosetothecommonancestorof6BanfieldJF,MarshallCR.Genomicsandthegeosciences.Sci-tetrapodsandlungfish.Nature,2002,418:767—770ence,2000,287:605—60628ShuD,ConwayMorrisS,ZhangZJ,etal.AnewspeciesofYun-7CutterGA.Biogeochemistry:Nowandintothefuture.Palaeo-nanozoanwithimplicationsforDeuterostomeevolution.Science,geogrPalaeoclimatolPalaeoecol,2005,219:191—1982003,299:1380—13848NaylorLA.Thecontributionsofbiogeomorphologytothe29DongX,DonoghuePCJ,ChengH,etal.Fossilembryosfromtheemergingfieldofgeobiology.PalaeogeogrPalaeoclimatolPa-MiddleandLateCambrianperiodofHunan,southChina.Nature,laeoecol,2005,219:35—512004,427:237—2409HuttonJ.Aninvestigationofthelawsobservableinthecomposi-30ChenJ,BottjerDJ,OliveriP,etal.Smallbilaterianfossils40totion,dissolutionandrestorationofLandupontheGlobe.TransR55millionyearsbeforetheCambrian.Science,2004,305:218—SocEdinb,1788,1(Ⅱ):413—42922210BassBeckingLGM.Geobiologieofinleidingtotdemilieukunde.31WangY,HuY,MengJ,etal.AfossifiedMeckel’scartilageinTheHague:WPVanStockum&Zoon.1934twoCretaceousmammalsandoriginofthemammalianmiddleear.11LovelockJE.Geophysiology:Anewlookatearthscience.BullScience,2001,294:357—361AmMeteorologSoc,1986,67:392—39732SunG,QiangJ,DilcherDL,etal.Archaefructaceae,anewbasal12CommitteeonFacilitatingInterdisciplinaryResearch,Committeeangiospermfamily.Science,2002,296:899—904onScience,EngineeringandPublicPolicy.FacilitatingInterdisci-33LuoZ,JiQ,WibleJR,etal.AnEarlyCretaceoustribosphenicplinaryResearch.WashingtonDC:TheNationalAcademiesPress,mammalandmetatherianevolution.Science,2003,302:1934—2004.142―143194013陈宜瑜,陈泮勤,葛全胜,等.全球变化研究进展与展望.地学34XuX,ZhouZ,WangX,etal.Four-wingeddinosaursfromChina.2334www.scichina.com\n论坛第51卷第19期2006年10月Nature,2003,421:335—34056StevensTO,McKinleyJP.Lithotrophicmicrobialecosystemsin35ZhouZ,ZhengS.ThemissinglinkinGinkgoevolution.Nature,deepbasaltaquifers.Science,1995,270:450—4542003,423:821—82257RasmussenB.Filamentousmicrofossilsina3235-million-year-old36KienastM,SteinkeS,StatteggerK,etal.Synchronoustropicalvolcanogenicmassivesulphide.Nature,2000,405:676—679southChinaSeaSSTchangeandGreenlandwarmingduringde-58AllwoodAC,WalterMR,KamberBS,etal.Stromatolitereefglaciation.Science,2001,291:2132—2134fromtheEarlyArchaeaneraofAustralia.Nature,2006,441:71437HinrichsK,HmeloLR,SylvaSP.Molecularfossilrecordofele-—718vatedmethanelevelsinlatePleistocenecoastalwaters.Science,59ShenY,BuickR,CanfieldDE.Isotopicevidenceformicrobial2003,299:1214—1217sulphatereductionintheearlyArchaeanera.Nature,2001,410:38JenkynsHC,ForsterA,SchoutenS,etal.Hightemperaturesin77—81theLateCretaceousArcticOcean.Nature,2004,432:888—89260KnollAH.Thegeologicalconsequencesofevolution.Geobiol-39HuangY,Street-PerrottFA,MetcalfeSE,etal.Climatechangeogy,2003,1:3—14asthedominantcontrolonglacial-interglacialvariationsinC3and61FalkowskiPG,KatzME,KnollAH,etal.Theevolutionofmod-C4plantabundance.Science,2001,293:1647—1651erneukaryoticphytoplankton.Science,2004,305:35436040XieS,YaoT,KangS,etal.GeochemicalanalysisofaHimalayan62SheehanPM,HarrisMT.MicrobialiteresurgenceaftertheLatesnowpitprofile:implicationforatmosphericpollutionandclimate.Ordovicianextinction.Nature,2004,430:75—78OrgGeochem,2000,31:15—2363LehrmannDJ,PayneJL,FelixSV,etal.Permian-Triassic41ZhangZ,ZhaoM,LuH,etal.Lowertemperatureasthemainboundarysectionsfromshallow-marinecarbonateplatformsofthecauseofC4plantdeclinesduringtheglacialperiodsontheChi-NanpanjiangBasin,southChina:Implicationsforoceaniccondi-neseLoessPlateau.EarthPlanetSciLett,2003,214:467—481tionsassociatedwiththeend-Permianextinctionanditsaftermath.42XieS,YiY,HuangJ,etal.LipiddistributioninasubtropicalPalaios,2003,18:138—152southernChinastalagmiteasarecordofsoilecosystemresponse64PrussSB,CorsettiFA,BottjerDJ.Theunusualsedimentarytopaleoclimatechange.QuatRes,2003,60:340—347rockrecordoftheEarlyTriassic:Acasestudyfromthesouth-43SachsJP,LehmanSJ.SubtropicalNorthAtlantictemperatureswesternUnitedStates.PalaeogeogrPalaeoclimatolPalaeoecol,60,000to30,000yearsago.Science,1999,286:756—7592005,222:33—5244SachsJP,AndersonRF,LehmanSJ.Glacialsurfacetemperatures65StephensNP,SumnerDY.Famennianmicrobialreeffacies,ofthesoutheastAtlanticOcean.Science,2001,293:2077—2079NapierandOscarRanges,CanningBasin,westernAustralia.45VenkatesanMI,DahlJ.OrganicgeochemicalevidenceforglobalSedimentology,2003,50:1283—1302firesattheCretaceous/Tertiaryboundary.Nature,1989,338:57—66RidingR,LiangL.Geobiologyofmicrobialcarbonates:metazoan60andseawatersaturationstateinfluencesonseculartrendsduring46EganhouseRP.MolecularMarkersinEnvironmentalGeochemis-thePhanerozoic.PalaeogeogrPalaeoclimatolPalaeoecol,2005,try.WashingtonDC:AmericanChemicalSociety,1997.1—415219:101—11547EhrlichHI.Geomicrobiology.NewYork:MarcelDekker,200267MiltnerA,RichnowH,KopinkeF,etal.AssimilationsofCO2by48BanfieldJF,NealsonKH.Geomicrobiology:InteractionsBe-soilmicroorganismsandtransformationintosoilorganicmatter.tweenMicrobesandMinerals.Reviewinmineralogy.WashingtonOrgGeochem,2004,35:1015—1024DC:MineralogicalSocietyofAmerica,199768HallettTB,CoulsonT,PilkingtonJG,etal.Whylarge-scalecli-49OrphanVJ,HouseCH,HinrichsK,etal.Methane-consumingmateindicesseemtopredictecologicalprocessesbetterthanlocalarchaearevealedbydirectlycoupledisotopicandphylogeneticweather.Nature,2004,430:71—75analysis.Science,2001,293:484—48769JennerjahnTC,IttekkotV,ArzHW,etal.Asynchronousterres-1450PetschST,EglintonTI,EdwardsKJ.C-deadlivingbiomass:trialandmarinesignalsofclimatechangeduringHeinrichevents.evidenceformicrobialassimilationofancientorganiccarbondur-Science,2004,306:2236—2239ingshaleweathering.Science,2001,292:1127—113170HughenKA,EglintonTI,XuL,etal.Abrupttropicalvegetation51KastingJF,SiefertJL.LifeandtheevolutionofEarth’satmos-responsetorapidclimatechanges.Science,2004,304:1955—phere.Science,2002,296:1066—1068195952TorsvikV,ØvreåsL,ThingstadTF.Prokaryoticdiversity——71FredricksonJK,PhelpsTJ.Subsurfacedrillingandsampling.In:magnitude,dynamicsandcontrollingfactors.Science,2002,296:KnudsenG,StetzenbachL,McInerneyMM,etal.,eds.Manualof1064—1066EnvironmentalMicrobiology.WashingtonDC:ASMPress,1996.53MichaelisW,SeifertR,NauhausK,etal.Microbialreefsinthe526—540BlackSeafueledbyanaerobicoxidationofmethane.Science,72EdwardsKJ,BondPL,GihringTM,etal.Anarchaeal2002,297:1013—1015iron-oxidizingextremeacidophileimportantinacidminedrainage.54KrumholzLR.Microbialcommunitiesinthedeepsubsurface.Science,2000,287:1796—1799HydrogeolJ,2000,8:41073WalkerJJ,SpearJR,PaceNR,etal.Geobiologyofamicrobial55XieS,PancostRD,YinH,etal.TwoepisodesofmicrobialendolithiccommunityintheYellowstonegeothermalenvironment.changecoupledwithPermo/Triassicfaunalmassextinction.Na-Nature,2005,434:1011—1014ture,2005,434:494—49774PriscuJC,AdamsEE,LyonsWB,etal.Geomicrobiologyofwww.scichina.com2335\n第51卷第19期2006年10月论坛subglacialiceaboveLakeVostok,Antarctica.Science,1999,286:95VisscherPT,StolzJF.Microbialmatsasbioreactors:populations,2141—2144processes,andproducts.PalaeogeogrPalaeoclimatolPalaeoecol,75KarlDM,BirdDF,BjörkmanK,etal.Microorganismsintheac-2005,219:87—100cretediceofLakeVostok,Antarctica.Science,1999,286:2144—96MarkošA.TheontogenyofGaia:theroleofmicroorganismsin2147planetaryinformationnetwork.JTheorBiol,1995,176:175—18076ReysenbachA,ShockE.Merginggenomeswithgeochemistryin97BrehmaU,GorbushinaA,MottersheadD.Theroleofmicroor-hydrothermalecosystems.Science,2002,296:1077—1082ganismsandbiofilmsinthebreakdownanddissolutionofquartz77ReysenbachA-L,LiuY,BantaAB,etal.Aubiquitousthermoaci-andglass.PalaeogeogrPalaeoclimatolPalaeoecol,2005,219:117dophilicarchaeonfromdeep-seahydrothermalvents.Nature,2006,—129442:444—44798DechoA.Exopolymermicrodomainsasastructuringagentfor78FiskMR,GiovannoniSJ,ThorsethIH.Alterationofoceanicheterogeneitywithinmicrobialbiofilms.In:RidingR,AwramikSvolcanicglass:texturalevidenceofmicrobialactivity.Science,M,eds.MicrobialSediments.Berlin:Springer,2000.9—151998,281:978—98079SharmaA,ScottJH,CodyGD,etal.Microbialactivityatgiga-99ArpG,ReimerA,ReitnerJ.Photosynthesis-inducedbiofilmcal-pascalpressures.Science,2002,295:1514—1516cificationandcalciumconcentrationsinphanerozoicoceans.Sci-80CowenJP,GiovannoniSJ,KenigF,etal.Fluidsfromagingence,2001,292:1701—1704oceancrustthatsupportmicrobiallife.Science,2003,299:120—100AndersonDL.TheEarthasaplanet:Paradigmsandparadoxes.123Science,1984,223:347—35481ZhangG,DongH,XuZ,etal.Microbialdiversityinul-101KrumbeinWE,StalLJ.Geophysiologyofmarinecyanobacterialtra-high-pressurerocksandfluidsfromtheChineseContinentalmatsandbiofilms.KielerMeeresforschungen,1991,8:137—145ScientificDrillingProjectinChina.AppEnvironMicrobiol,2005,102GrimmKA,LangeCB,GillAS.Self-sedimentationofphyto-71:3213—3227planktonbloomsinthegeologicrecord.SedimentGeol,1997,110:82BräuerSL,Cadillo-QuirozH,YashiroE,etal.Isolationofa151—161novelacidiphilicmethanogenfromanacidicpeatbog.Nature,103RameyBE,KoutsoudisM,vonBodmanSB,etal.Biofilmfor-2006,442:192194mationinplant-microbeassociations.CurrOpinMicrobiol,2004,83TrollC.Landscapeecology(geoecology)andbiogeocenology-A7:602—609terminologicalstudy.Geoforum,1971,8:4346104FuxCA,CostertonJW,StewartPS,etal.Survivalstrategiesof84HuggettRJ.Geoecology:AnEvolutionaryApproach.London:infectiousbiofilms.TrendsMicrobiol,2005,13:34—40Routledge,1995.320105JinYG,WangY,WangW,etal.Patternofmarinemassextinction85TrofimovVT.Ecologicalgeology-anovelbranchofgeologicalnearthePermian-TriassicboundaryinSouthChina.Science,2000,sciences.EarthSciFront,2001,8:27—35289:432—43686LovelockJE.Geophysiology,thescienceofGaia.RevGeophys,1989,27:215—222106KelleyPH,KowalewskiM,HansenTA.Predator-PreyInterac-87LovelockJE,KumpLR.Failureofclimateregulationinageo-tionsintheFossilRecord,TopicsinGeobiologySeries.NewYork:physiologicalmodel.Nature,1994,369:732—734Kluwercademic/PlenumPublishers,2003.1—46488McMenaminMAS,McMenaminDLS.Hypersea:LifeonLand.107周忠和,骆永明,董云社,等.浅谈中国地球环境与生命过程研NewYork:ColumbiaUniPress,1996究中的一些科学问题.科学通报,2005,50(2):195—20089VolkT.Gaia’sBody:TowardsaPhysiologyofEarth.NewYork:108汪品先.中国的地球系统科学研究向何处去.地球科学进展,Copernicus/SpringerVerlag,19982003,18(6):837—85190张弥曼,陈旭.20世纪古生物学的重大进展及21世纪战略重点.109PapidaS,MurphyW,MayE.Enhancementofphysicalweather-地球科学进展,2001,16(5):624—628ingofbuildingstonesbymicrobialpopulations.IntBiodeterior91SterflingerK.Fungiasgeologicalagents.GeomicrobiolJ,2000,Biodegrad,2000,46:305—31717:97—124110GorbushinaAA,KrumbeinWE,VolkmannM.Rocksurfacesas92BurfordEP,FominaM,GaddGM.Fungalinvolvementinbiow-lifeindicators:newwaystodemonstratelifeandtracesofformereatheringandbiotransformationofrocksandminerals.Minerallife.Astrobiology,2002,2:203—213Mag,2003,67:1127—1155111TsudaA,TakedaS,SaitoH,etal.Amesoscaleironenrichmentin93BernerRA.Thelong-termcarboncycle,fossilfuelsandatmos-theWesternSubarcticPacificinducesalargecentricdiatomphericcomposition.Nature,2003,426:323—326bloom.Science,2003,300:958—96194GorbushinaAA,BoettcherM,BrumsackH-J,etal.Biogenic112汪品先.探索大洋碳储库的演变周期.科学通报,2003,48(21):forsteriteandopalasaproductofbiodeteriorationandLichen2216—2227Stromatoliteformationintablemountainsystems(Tepuis)ofVenezuela.GeomicrobiolJ,2001,18:117—132(2006-04-29收稿,2006-08-16接受)2336www.scichina.com查看更多