- 2022-08-12 发布 |
- 37.5 KB |
- 7页


申明敬告: 本站不保证该用户上传的文档完整性,不预览、不比对内容而直接下载产生的反悔问题本站不予受理。
文档介绍
发育生物学考试习题
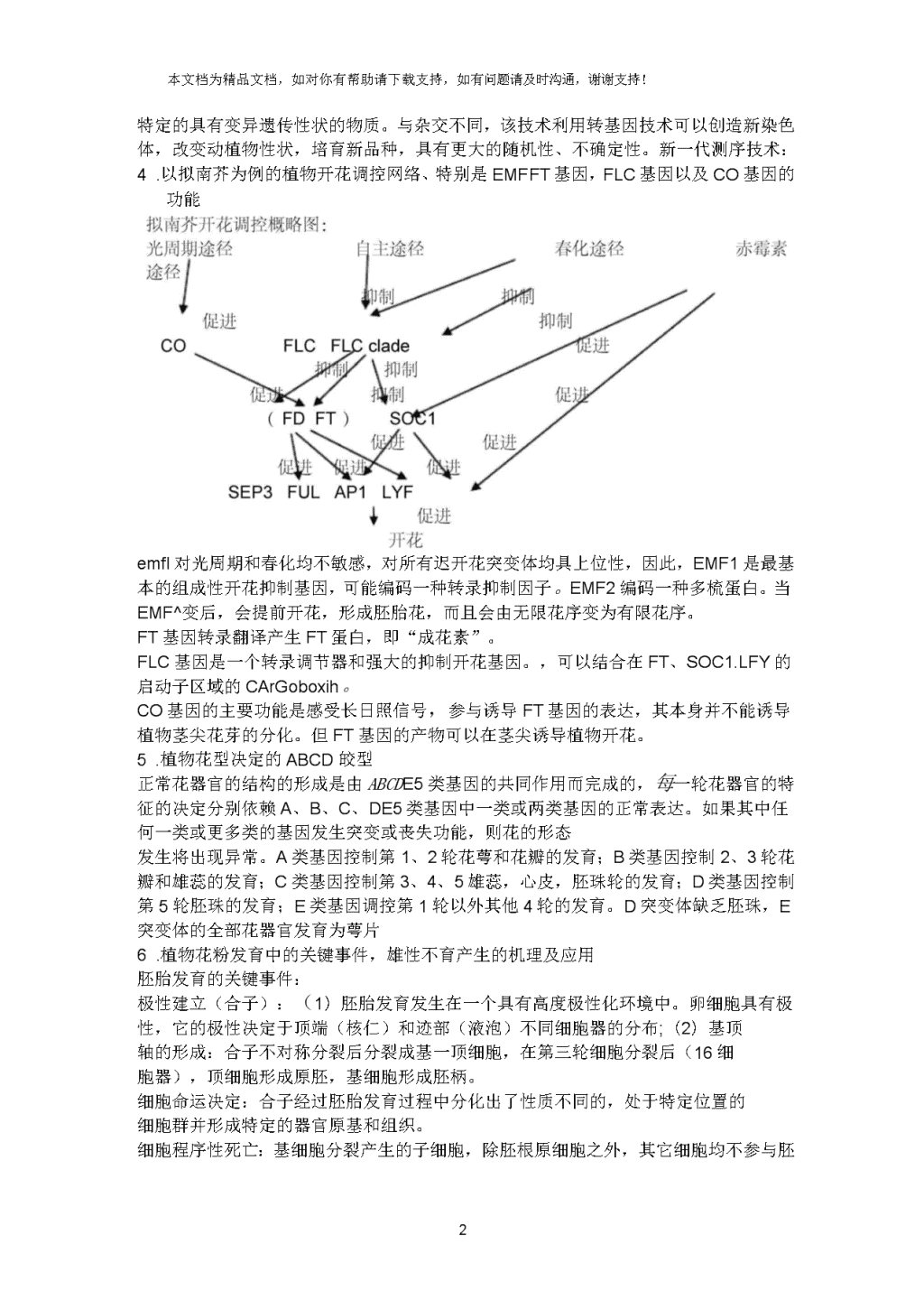
本文档为精品文档,如对你有帮助请下载支持,如有问题请及时沟通,谢谢支持!重点内容1.动植物发育的异同点植物是一种形态学上简单的有机体,动物的组成则比较复杂;植物细胞具有全能性,动物细胞则不具有;植物既可以用抱子,也可以用配子进行生殖,动物只能形成配子进行生殖;高等植物中具有双受精现象,动物中则不存在这种情况;植物胚胎发生完成于休眠种子的形成,而动物则与之既然不同动物的胚胎发生没有休眠过程。;高等植物的胚形成两个具有不同发育命运的器官系统,动物则没有;植物具有无限的发育程序,动物则没有;植物的形态建成在没有细胞移动的情况下进行,动物细胞在胚胎发育中可移动位置;环境因子在植物发育中扮演主要的角色,动物则在环境中可自由移动,有一定逃避不良环境的能力。2.模式动物、模式植物应该具备哪些特性其生理特征能够代表生物界某一大类群;容易获得并易于在实验室内饲养,繁殖;容易进行试实验操作,特别是遗传分析;生命结构都很简单。个体小,生育期短,生殖能力强、染色体组成简单,染色体数目少,基因组少,且测序完成3.发育生物学研究的主要方法:显微观察、细胞谱系追踪、突变体构建、图位克隆、基因表达分析、转基因、新一代测序技术生物统计和生物信息学,表达谱构建和分析显微观察:使用显微设备进行细胞、组织、结构研究。细胞谱系追踪:细胞间在发育中世代相承的亲缘关系犹如人类家族的谱系称为“细胞谱系”。随着现代分子生物学技术发展,一些新的技术也运用到细胞谱系的研究中去。例如:基因陷阱是一种基于报告基因表达的方法,用在某些器官特异表达的或者35S启动子连接GU邻报告基因来研究细胞谱系,也有人用X射线照射来研究细胞谱系,荧光原位杂交(florescencein-situhybridization,FISH)是一种非放射性原位杂交方法。突变体构建:(重要且基本的实验材料)利用自然发生或人工诱导发生的突变体研究突变基因的结构及其在植物体发育过程中的功能和调控等。图位克隆:又称定位克隆,是新型分离和克隆植物基因的方法之一,在不知道基因的表达产物,在未知基因的功能信息又无适宜的相对表型用于表型克隆时,最常用的基因克隆技术就是图位克隆法。图位克隆是通过分析突变位点与已知分子标记的连锁关系来确定突变表型的遗传基本。原理:用该方法分离基因是根据功能基因在基因组中都有相对稳定的基因座,再利用分子标记技术对目的基因进行精确定位的基础上,用与目的基因紧密连锁的分子标记筛选DNAC库,从而构建目的基因区域的物理图谱,再利用此物理图谱通过染色体步移逐步逼近目的基因或通过染色体登陆的方法,最终克隆目的基因并通过遗传转化实验可以研究目的基因的功能。基因表达分析:是通过快速和详细分析成千上万个EST(expresssequencedtags)来寻找出表达丰富度不同的SAG所签序列。在此方法中,通过限制性酶切可以产生非常短的cDNA(10-14bp)标签,并通过PCRT增和连接,随后对连接体进行测序。转基因:是指运用科学手段从某种生物体中提取所需要的基因,将其转入另一种生物中,使与另一种生物的基因进行重组,再从结果中进行数代的人工选育,从7\n本文档为精品文档,如对你有帮助请下载支持,如有问题请及时沟通,谢谢支持!而获得特定的具有变异遗传性状的物质。与杂交不同,该技术利用转基因技术可以创造新染色体,改变动植物性状,培育新品种,具有更大的随机性、不确定性。新一代测序技术:1.以拟南芥为例的植物开花调控网络、特别是EMFFT基因,FLC基因以及CO基因的功能emfl对光周期和春化均不敏感,对所有迟开花突变体均具上位性,因此,EMF1是最基本的组成性开花抑制基因,可能编码一种转录抑制因子。EMF2编码一种多梳蛋白。当EMF^变后,会提前开花,形成胚胎花,而且会由无限花序变为有限花序。FT基因转录翻译产生FT蛋白,即“成花素”。FLC基因是一个转录调节器和强大的抑制开花基因。,可以结合在FT、SOC1.LFY的启动子区域的CArGoboxih。CO基因的主要功能是感受长日照信号,参与诱导FT基因的表达,其本身并不能诱导植物茎尖花芽的分化。但FT基因的产物可以在茎尖诱导植物开花。2.植物花型决定的ABCD皎型正常花器官的结构的形成是由ABCDE5类基因的共同作用而完成的,每一轮花器官的特征的决定分别依赖A、B、C、DE5类基因中一类或两类基因的正常表达。如果其中任何一类或更多类的基因发生突变或丧失功能,则花的形态发生将出现异常。A类基因控制第1、2轮花萼和花瓣的发育;B类基因控制2、3轮花瓣和雄蕊的发育;C类基因控制第3、4、5雄蕊,心皮,胚珠轮的发育;D类基因控制第5轮胚珠的发育;E类基因调控第1轮以外其他4轮的发育。D突变体缺乏胚珠,E突变体的全部花器官发育为萼片3.植物花粉发育中的关键事件,雄性不育产生的机理及应用胚胎发育的关键事件:极性建立(合子):(1)胚胎发育发生在一个具有高度极性化环境中。卵细胞具有极性,它的极性决定于顶端(核仁)和迹部(液泡)不同细胞器的分布;(2)基顶轴的形成:合子不对称分裂后分裂成基一顶细胞,在第三轮细胞分裂后(16细胞器),顶细胞形成原胚,基细胞形成胚柄。细胞命运决定:合子经过胚胎发育过程中分化出了性质不同的,处于特定位置的细胞群并形成特定的器官原基和组织。细胞程序性死亡:基细胞分裂产生的子细胞,除胚根原细胞之外,其它细胞均不7\n本文档为精品文档,如对你有帮助请下载支持,如有问题请及时沟通,谢谢支持!参与胚的形成,而是共同构成胚柄,将胚胎与珠孔端的母体组织连在一起,并在心形胚后期开始进入程序化衰老而退化。基因表达调控:(1)基顶轴极性中wox2/wox8,9调控途径;(2)外层细胞(AtMLI和ACR妙同决定)和内层细胞(MPTF口MO7调控基因。花粉发育的关键事件:从造抱细胞一小抱子母细胞叫分体f抱子花粉主要是两个阶段,阶段一:小抱子发生,第二个阶段:花粉粒形成并释放。这两个阶段主要发生了:小抱子经一次不对称的有丝分裂产生一个营养核一个精核,精细胞再经过一次有丝分裂形成两个精子雄性不育产生的机理及应用:雄性不育:植株由于不能产生正常的花药、花粉或雄配子而导致不育的现象。主要类型:细胞质雄性不育(玉米的T型胞质)。细胞核不育:显性核不育隐性核不育。应用•:利用雄性不育进行杂交育种,3系选育1.植物自交不亲和性类型及决定方式(重点掌握十字花科与蔷薇科)配子体型不亲和(gamatophyticself■Iincompatibility,GSI,即受花粉本身的基因控制)。GSI发生在花柱中,表现为花粉管生长停顿、破裂;抱子体型不亲和(sporphyricself巾incompatibilitySSI,即受花粉亲本基因控制)。SSI发生于柱头表面,表现为花粉管不能穿过柱头。2.拟南芥(或者芹菜)胚胎发育的一般模式及关键事件;胚乳发育中的基因组印迹拟南芥(或者芹菜)胚胎发育的一般模式:伸长期合子;合子经过第一次分裂,产生1个顶细胞(绿色)和一个基细胞(粉红色);四分体胚;16细胞球形胚;早期心形胚;心形胚;成熟胚。关键事件:极性建立(合子):(1)胚胎发育发生在一个具有高度极性化环境中。卵细胞具有极性,它的极性决定于顶端(核仁)和迹部(液泡)不同细胞器的分布;(2)基顶轴的形成:合子不对称分裂后分裂成基一顶细胞,在第三轮细胞分裂后(16细胞器),顶细胞形成原胚,基细胞形成胚柄。细胞命运决定:合子经过胚胎发育过程中分化出了性质不同的,处于特定位置的细胞群并形成特定的器官原基和组织。细胞程序性死亡:基细胞分裂产生的子细胞,除胚根原细胞之外,其它细胞均不参与胚的形成,而是共同构成胚柄,将胚胎与珠孔端的母体组织连在一起,并在心形胚后期开始进入程序化衰老而退化。基因表达调控:(1)基顶轴极性中wox2/wox8,9调控途径;(2)外层细胞(AtML1和ACR妙同决定)和内层细胞(MPTf口MO7调控基因。胚乳发育中的基因组印迹:是一种非孟德尔遗传现象,它是指在配子,合子发生期间来自亲本的等位基因或染色体产生专一性的加工修饰导致后代细胞中两个亲本来源的等位基因有不同的表达活性,又称遗传印记或亲代印记或配子印记。3.根尖及茎尖发育调控机制及异同点,SAMWRAM植物根的发育(主根,侧根与根毛)根尖及茎尖发育调控机制及异同点,SAMWRAIM植物根的发育(主根,侧根与根毛):植物在根部不同的位置产生不同的侧生根,主根在胚胎的下胚轴的基部就已经形成,然后在主根上形成侧根,但根尖7\n本文档为精品文档,如对你有帮助请下载支持,如有问题请及时沟通,谢谢支持!并不形成侧根。主根的中拄鞘侧向生长形成侧根。在根的某些地方形成根毛,但并不形成中柱鞘。侧根在预期的地方膨胀,侧根通常与木质部毗连。侧根继续膨胀,穿过内皮层,表皮形成侧根。在根部分化区,表皮细胞伸长产生根毛,在细胞达到分化区之前,细胞开始膨胀,当根毛长到一定长度以后,表皮细胞停止继续伸长,根中分化为根毛得细胞称为毛状细胞。1.植物幼苗光形态建成的分子机制当光被植物体吸收后,光感受器将与别的信号传导因子相互作用,这可以引起许多分子和形态学上的反应。植物通过光合作用将光能转化成化学能储存起来,光还能以信息的形式作用于植物从而调节植物的分化、生长、发育,使其能够更好地适应外界环境。这种调节通过生物膜系统结构、透性的变化和基因表达的变化促成了细胞的分化及其结构和功能的改变,最终汇集成组织和器官的建成,这就是光形态建成(photomorphogenesis),亦即光调控发育过程。从表型上看,植物在光下的形态结构变化主要表现在4个方面:下胚轴不伸长(hypocotel),子叶伸展(cotyledon),去白化(de-eti01ate),SAMFF始激活。分子机制:植物体内存在光受体,它能识别光信号并接受光信号,然后通过一系列的光信号传导途径将光信号传导到植物体内。然后调控植物机体从而表现出以上我们描述的形态上的变化。目前存在的光受体主要有光敏色素(phytochromes),隐花色素(cryphtochrome)和趋光性(phototropin)三种光受体。光敏素存在两种形式,且随吸收红光或远红光而互相转化一一活化态(Pf)r或钝化态(Pr),当其处于活化态时才能起作用。高等植物体内存在两种类型的光敏色素分子,即光敏素I(phyl)和光敏素H(phyH)。phyI是一种易溶于水的浅蓝绿色蛋白质,以二聚体形式存在于细胞质中;phyn主要存在于绿色组织中,吸收红光(R)转换为活化态(Pfr)后稳定存在,且在光下、暗中均可合成,phyII也是以二聚体的形式存在。phyI类光敏素,主要接收波长700〜750nm的远红光;phyII类光敏素,主要接收波长600〜700nm的红光。隐花色素(cryptochrome)在发育过程中则可以调控植物的伸长生长,开花时间以及生物体的生理节奏。隐花色素包括功能相似的CRY便口CRY2W种类型,其中CRY2勺C-端较CRY1的短。而它们却都可以阻止下胚轴伸长、诱导CHSS因表达、促进花青素合成,并可以吸收将光敏色素吸收的红光。第三种类型是趋光性受体(丝氨酸/色氨酸激酶),这中类型的光受体可以调控植物的一系列运动反应,包括趋光性、叶绿体运动及气孔开启。到目前为止对于这一对激酶人们已经在植物中发现了两种即Phot1和Phot2。2.植物侧生器官的形成,叶片发育中的极性形成,侧枝发育的激素调控;株高调控;理想株型植物侧生器官的形成:SAM可以产生一系列的植物营养体单位,由叶、腋生SAM和节间组成,最终发育成一些侧生器官,包括叶,侧枝等。叶形成于茎顶端分生组织SAM产生的叶原基侧枝形成过程:SAM^生月S生SAM腋生SAMHt产生侧芽原基,侧芽原基生长为侧芽,侧芽继续发育成侧枝。叶片发育中的极性形成:叶片的极性包括以下3方面:1、近-远轴:从叶的上表面指向下表面;PHB/PHV选择性的在近轴面表达是叶片近-远轴极性获得的关键。2、中-边轴:从叶的主脉指向边缘;先中脉后叶片,从中肋向边缘。3、基-顶轴:由叶的基部指向尖部7\n本文档为精品文档,如对你有帮助请下载支持,如有问题请及时沟通,谢谢支持!KN1影响基-顶轴形成,显性的kn1突变会导致沿着叶片侧生脉的组织形成结,原因:叶的基-顶轴可能是由SAM产生的一种化学彳S号所决定的。KN1蛋白可能会诱导这种形态发生素的产生侧枝发育的激素调控:侧芽的形成与侧枝的发育是细胞分裂素与生长素两种激素综合作用的。植物顶端产生的生长素可以极性向下运输,并进入侧芽,从而直接抑制侧芽的生长,MAXsB因通过抑制生长素的运输载体PIN蛋白的活性,使侧芽中的生长素无法向外转运,从而抑制侧芽伸长。细胞分裂素是侧芽生长的一个决定因素,促进侧芽生长伸长。生长素/细胞分裂素比值高有利于主枝生长,根伸长生长,生长素/细胞分裂素比值低促进侧枝发育、侧根增多株高调控:激素与株高:赤霉素(Gibberellins,GAs)油菜素内酯(Brasinosteriod,BR)生长素(Indole-3-aceticacid,IAA)一些同源异型盒基因、转录因子、细胞壁形成基因也会影响植物的株在拟南芥中,控制株高的GA1基因,即赤霉素合成酶基因,突变后ga1,植株矮小,SPYS因,编码GAS应蛋白,抑制GA勺信号传递,突变后,会使植株变高。水稻中的“绿色革命基因sd-1,植株矮,水稻中克隆了水稻长节间基因ELONGATEDUPPERMOSTINTERN!E编码一种细胞色素P450monooxygenaseCYP714D1与GA合成有关,突变后植株变高BUI1基因:BUI1编码一个植物特异的ClassHformin蛋白,调控细胞微丝骨架的装配和动态变化。突变后,变矮。玉米中大约有50种基因影响株高,zong3,变矮。Zong3的株高较低主要由细胞长度短造成油菜ds-1基因,该基因为BnRGA编码一个DELLAg白,参与赤霉素信号转导,该基因为单碱基变异,不影响基因表达量,突变后功能加强,进一步抑制了GA信号传导,植株变矮理想株型:1.拟南芥角果成熟的一般过程,种子散发调控机理;西红柿果实大小的决定拟南芥角果成熟的一般过程:1.花原基起始2.花原基隆起并与SAMSES3.花萼原基形成4.花萼近远轴形成5.花瓣与雄蕊原基产生6.萼片包围花芽及雌蕊形成(顶端细胞不分裂,导致凹陷形成两侧发育,形成中空的管状结构)7.中间花药突起及中空状雌蕊形成8.花药中小室形成及管状雌蕊增大9.花瓣原基产生及隔膜形成11.花瓣原基达到较短的花药的高度12.柱头乳突细胞形成,内外珠被开始形成7\n本文档为精品文档,如对你有帮助请下载支持,如有问题请及时沟通,谢谢支持!11.花瓣达到较长花药高度,雌蕊成熟,花柱形成12.花开放,传粉、授精13.花药达到了柱头的高度、授精完成14.雌蕊进一步生长15.花瓣、萼片、雄蕊凋谢16.角果发育17.果实成熟及开裂种子散发调控机理:决定种子散发的基因调控:SHPIND,ALGRPLFULSHP对木质化层和分离层的形成具有重要作用。IND专一化的决定木质化层的形成。ALC影响分离层分化。RPLFUL西红柿果实大小的决定:细胞分裂速度和小室数。13.水稻产量形成的主要因素及主要决定基因主要因素:单株穗数;每穗粒数;种子大小;主要决定基因:moc1定位于细胞核中,主要在腋芽处表达,促进腋芽起始和生长;HTD1该基因突变后导致分奠增多,株高变矮,解除了对侧芽的抑制所致。摘除侧芽后可以部分恢复株高表型;D10控制分裂数;LAX基因突变导致花序轴,分枝数,小穗数均受到影响(LAX与SPA<突变体可以加强表型,几乎没有分枝和小穗形成);FZP控制水稻花型;FZP或者抑制小穗原基想侧芽原基转变,或者促进其向花原基转变;籽粒大小决定-GS3-粒长;籽粒大小决定-GW2粒宽。14.PCD在植物发育中的重要作用及其发生机理;植物衰老的类型及发生过程细胞程序性死亡(PCD)是细胞主动的有序的死亡。它主要表现在细胞萎缩、核固缩、染色质凝集、细胞形成“凋亡小体”和“凋亡小体”被吞噬等。作用:在生殖器官的发育中保证功能细胞的发育和生殖过程的完成;在胚胎发育中保证受精卵发成正常胚胎;在种子萌发中保证幼苗的形成;在植物发育中保证有关器官的建成和组织分化;在免疫反应和抗病中的作用。发生机理:植物衰老的类型及发生过程:7\n本文档为精品文档,如对你有帮助请下载支持,如有问题请及时沟通,谢谢支持!衰老是由基因控制的且受内外因素影响的导致植物自然死亡的一系列衰退过程。类型:整体衰老;地上部分衰老;脱落衰老;渐进衰老;过程:13.试卷论述题第二题我们知道,自上个世纪中叶分子生物学迅速发展以来,为生物科学的研究方法带来了革命性的改变,虽然植物分子生物学的研究曾经较动物和微生物相对滞后,但在近十余年来,随着现代生物技术方法的不断进步,植物发育生物学研究进展相当迅速。由于植物科学与农业生产实践息息相关,了解植物的生长发育机理和调控机制,对提高农作物产量,改善农作物品质具有重要意义,这在某种程度上也促进了植物发育生物学的发展。植物发育生物学的研究仍将从深度和广度两个方面发展。从深度来说,目前各国生物科学工作者们以分离到了一系列植物发育特定事件相关基因,并在细胞和分子水平上对其表达调控规律及其生物学功能进行了较深入透彻的分析。这些研究不仅在植物发育生物学理论上极为重要,而且也将为农业生产实践提供理论与技术指导。从广度来说,将拟南芥这种模式植物研究清楚后,应用于各种作物,改良作物的品质和产量,应用于生产,造福全人类是最终目的。另外,逆境胁迫的研究也需要深入。SAM:SAMfc胚发育的鱼雷期出现,SAM勺组成:是有三个相互联系的细胞层组成的,包含3个功能区域,中心区:由干细胞组成,周缘区:叶原基的起始位点.髓分生组织:茎髓的起始位点.Sam的发育:STM(SHOOTMERISTEMLESS)USCHEL(WUS)CLAVATA1(CLV1)CLAVATA3(CLV3)STM是胚胎中形成SA晰必需的WU编码一个同源异性域转录因子,在中心区表达,WUSt中心区表达,诱导CLV3在L1,L2层表达。产生的CLV3蛋白,激活L3中的CLV1信号途径;反之,CLV1的激活,又限制了WUS勺表达区域RAM:根顶端分生组织并不在根的最端,而是位于根冠保护性盖的后面.由胚的分生组织发育,由根冠、表皮、皮层、内皮层、维管束、不活动区组成,Ram3勺发育:WOX®因在RAM勺干细胞中起重要作用.SCR&QC内皮层、皮层初始区和已分化的内皮层表达,其功能缺失导致起始区及QC干细胞的连续性丧失;SHR&中柱细胞表达,其蛋白质转运到邻近细胞(包括QC).SHR功能丧失导致QC结构不规则,缺乏QC#异性和根部停止生长7查看更多